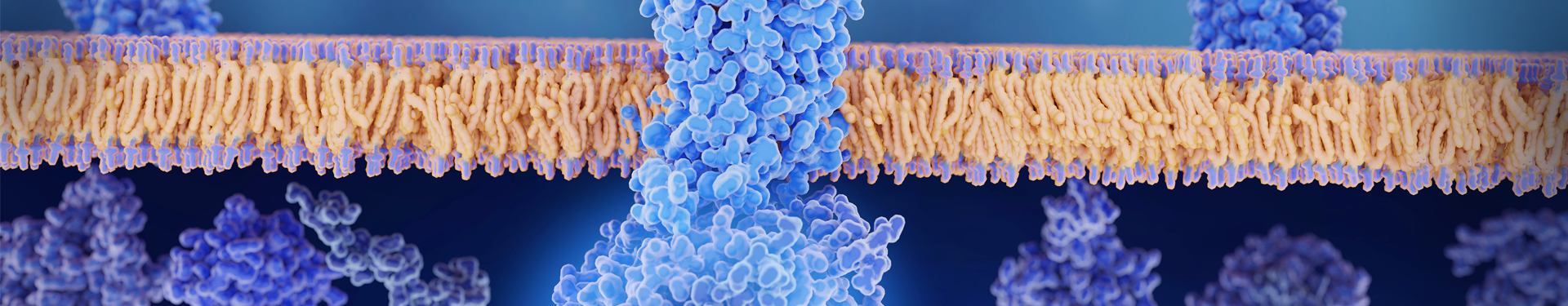
Gli RNA circolari (circRNA) furono scoperti per la prima volta come RNA non codificanti nel 1979; è stato solo nel 2015 che i circRNA trascritti sono stati trovati in Drosophila, il che ha portato allo studio dei circRNA in vitro per scopi terapeutici e preventivi.
I circRNA si autoespandono e quindi non hanno una struttura convenzionale come un cappuccio da 5' o una coda poliA da 3'. Questa struttura li rende più stabili e resistenti alle esonucleasi come la RNasi R.
Elementi di struttura dell'RNA circolare (circRNA)
Il DNA modello per la sintesi del circRNA include tipicamente il sito di ingresso ribosomiale interno (IRES), il frame di lettura aperto (ORF) e altri che sono fondamentali per la circolazione in vitro. Ad esempio, abbiamo progettato un plasmide come modello di trascrizione in vitro (IVT), che consiste di bracci di omologia, introne 3', esone 3', IRES, ORF, esone 5' e introne 5', per preparare il circRNA mediante auto-giunzione metodi.
Il sito di ingresso ribosomiale interno (IRES) svolge ruoli importanti nella traduzione dei cirRNA. L'IRES del virus dell'encefalomiocardite (EMCV), l'IRES del coxsackievirus B3 (CVB3), l'IRES del rinovirus umano B3 (HRV-B3) e altri IRES sono stati comunemente utilizzati per la traduzione della sintesi in vitro dei cirRNA
La ciclizzazione in vivo dei precursori lineari del circRNA fornisce un passo importante nella sintesi del circRNA, solitamente attraverso percorsi di autoreplicazione chimici, enzimatici e mediati dai ribosomi. Conosciuto come una pietra miliare nel settore, l'approccio chimico per la produzione del circRNA, introdotto nel 1988, non è più utilizzato oggi a causa del suo costo elevato, della bassa resa, dell'elevato numero di sottoprodotti e del fatto che è adatto solo per la ciclizzazione RNA lunghi fino a 70 nucleotidi.
I metodi enzimatici per ciclizzare l'RNA si basano sugli enzimi T4 o ribozimi del batteriofago come T4 DNA ligasi (T4 Dnl 1), T4 RNA ligasi 1 (T4 Rnl 1) e T4 RNA ligasi 2 (T4 Rnl 2). Per formare RNA circolare da parte degli enzimi del batteriofago T4, i monofosfati nucleosidici devono essere localizzati all'estremità 5' e coniugati al gruppo OH all'estremità 3' dell'RNA. Poiché durante la reazione vengono aggiunti nucleosidi trifosfati, l'RNA IVT contiene guanosina trifosfato (GTP) all'estremità 5'.
I ribozimi sono sequenze di RNA che promuovono l'auto-giunzione convertendo le molecole di RNA lineari in circRNA senza la necessità di enzimi aggiuntivi. Il processo di autoassemblaggio prevede due reazioni consecutive di transesterificazione in siti specifici per garantire la formazione dei prodotti cirRNA desiderati. Il metodo di auto-giunzione degli introni del gruppo I, noto anche come PIE (incapsulamento di introni ed esoni), che consente la produzione di cirRNA più lunghi di 5 kb, è stato ampiamente studiato e si è dimostrato utile.
Applicazione dell'RNA circolare (circRNA)
Vaccini CircRNA
Come gli mRNA lineari, i circRNA possono essere convertiti in determinate proteine nelle cellule bersaglio e indurre una robusta immunità umorale e cellulare. Recentemente, ci sono alcuni gruppi di ricerca che hanno sviluppato con successo vaccini circRNA per prevenire COVID-19. I loro risultati non solo hanno mostrato i vantaggi dei vaccini a mRNA lineare, ma hanno anche vantato una migliore stabilità e una durata più lunga dell’espressione proteica rispetto all’mRNA lineare. Pertanto, i vaccini circRNA possono indurre risposte immunitarie adeguate anche a basse dosi.
Inoltre, alcuni ricercatori ritengono che i vaccini circRNA, in quanto mRNA di prossima generazione, potrebbero diventare in futuro strumenti efficaci per combattere le comuni malattie virali/batteriche e le principali malattie infettive emergenti, nonché per il trattamento del cancro e di altre malattie.
CircRNA nella terapia CAR/TCR-T
In qualità di pioniere nel campo dell'RNA circolare, ORNA ha sviluppato una terapia con recettori dell'antigene chimerico (CAR) in situ che utilizza ORN-101, un RNA circolare incapsulato in LNP, per modulare le cellule immunitarie nei pazienti. ORN-101 presenta un'elevata espressione del CAR guidato da un elemento IRES ottimizzato. Nei modelli animali, è stato dimostrato che ORN-101 induce la soppressione e la distruzione del tumore, suggerendo che le terapie antitumorali basate su ORN-101 possono interferire con la terapia convenzionale con cellule CAR-T.
Inoltre, Zhang et al. valutato la vitalità e l'efficacia terapeutica dei circRNA nelle terapie T-T antigene-specifiche del recettore delle cellule T (TCR). Hanno progettato un circRNA che codifica pp65-TCR-T prendendo di mira l'epitopo pp65 del citomegalovirus (CMV). Inoltre, è stato dimostrato che pp65-TCR è espresso sulle cellule T primarie per oltre 7 giorni. Inoltre, le cellule circRNA-pp65-TCR-T hanno ucciso in modo specifico e coerente le cellule tumorali che esprimono pp65 e HLA e hanno prolungato significativamente il tempo di sopravvivenza dei topi.
È stato inoltre dimostrato che le cellule trasfettate con circRNA pp65 mostravano risposte immunitarie migliori rispetto all'mRNA lineare.
Soluzione CRDMO bio-farmaceutica One-Stop di Yaohai per RNA a codifica lunga
 EN
EN
 AR
AR
 HR
HR
 CS
CS
 DA
DA
 NL
NL
 FI
FI
 FR
FR
 DE
DE
 EL
EL
 IT
IT
 JA
JA
 KO
KO
 NO
NO
 PL
PL
 PT
PT
 RO
RO
 RU
RU
 ES
ES
 SV
SV
 IW
IW
 ID
ID
 LV
LV
 LT
LT
 SR
SR
 SK
SK
 SL
SL
 UK
UK
 VI
VI
 ET
ET
 HU
HU
 TH
TH
 TR
TR
 FA
FA
 AF
AF
 MS
MS
 BE
BE
 MK
MK
 UR
UR
 BN
BN